
Forschungsstand 2018
von Jan Sütterlin
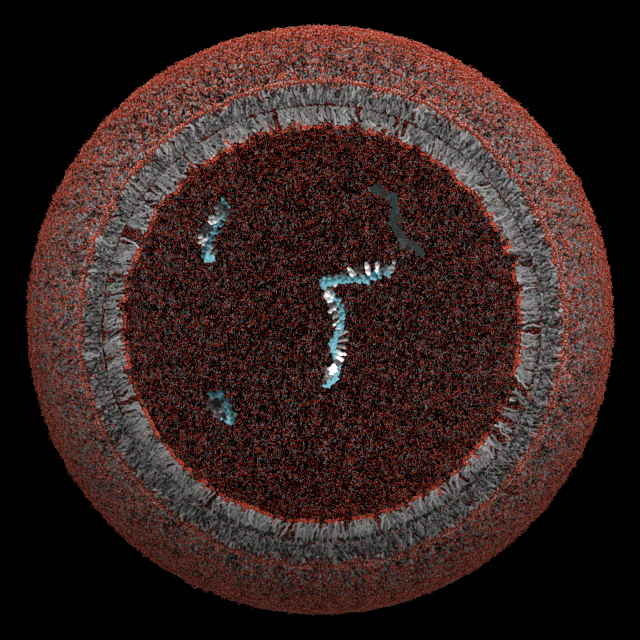
Titelbild: Querschnitt einer hypothetischen ersten Zelle (Protozelle). Illustration von Janet Iwasa, Lizenz CC BY-NC-ND 3.0 US.
Die Entstehung des Lebens bleibt eines der größten (noch) unbeantworteten Rätsel der Wissenschaft. Wie konnten sich die notwendigen chemischen Bausteine unter den Bedingungen der frühen Erde bilden und wie konnten sie sich zu einer ersten „lebenden“ Protozelle verbinden? Der Chemie kommt bei der Beantwortung dieser Frage eine Schlüsselrolle zu und hier konnten in den letzten 20 Jahren aufregende Erfolge erzielt werden: Ein Rückblick.
Seit der sowjetische Biochemiker Alexander Oparin 1924 in seinem Buch „Die Entstehung des Lebens auf der Erde“ die Hypothese aufstellte, dass wir nicht „von einem absoluten und grundsätzlichen Unterschied zwischen lebender und toter Materie sprechen müssen“ und, dass „Leben eine neue Form der Existenz der Materie ist, die unbedingt als eine bestimmte Etappe in der historischen Entwicklung von Materie auftreten musste“, versuchen Chemiker eine Brücke zur Biologie zu bauen. [1] [2] [3]
Um zu wissen wonach gesucht wird bedarf es zunächst einer Definition von Leben. Die gängige Definition aus biologischer Perspektive beinhaltet mindestens drei Kriterien: Reproduktion, Metabolismus und Mutation. [3] Über zusätzliche Kriterien besteht bis heute kein Konsens, es besteht jedoch der Verdacht, dass diese ohnehin aus den erstgenannten ableitbar sind. Nur durch Reproduktion gepaart mit gelegentlichen Veränderungen des Erbmaterials (Mutation) ist der Prozess der natürlichen Selektion vorstellbar. Ein Stoffwechsel (Metabolismus) ist notwendig um nachhaltige Wachstums-Reproduktionszyklen zu durchlaufen und einen geordneten, d.h. energiereichen, Zustand des lebenden Systems zu erhalten. Dies bedeutet, dass zumindest eine Aufnahme von Molekülen erfolgen muss, die in diesem Kontext als Nährstoffe bezeichnet werden können. Da Viren keinen eigenen Metabolismus aufweisen entsprechen sie dieser Definition von Leben nicht. Ohne den Metabolismus einer lebenden Wirtszelle auszunutzen können sie sich nicht reproduzieren.
Die Protozelle
Trotz der beeindruckenden makroskopischen Vielfalt sind sich alle Lebewesen, so wie wir sie auf der Erde kennen, auf mikroskopischer Ebene verblüffend ähnlich. Die kleinste funktionale Einheit aller Lebewesen ist die Zelle und die Erbinformation besteht aus nur wenigen identischen chemischen Bausteinen. Diese Tatsache weist stark auf gemeinsame Vorfahren hin. Diese ersten hypothetischen Lebewesen werden Protozellen genannt.
Wissenschaftler versuchen nun herauszufinden wie eine funktionierende Protozelle aufgebaut sein könnte, wie die einzelnen Bausteine zusammenfanden und wie diese Bausteine wiederum unter den primitiven Reaktionsbedingungen der Urerde entstanden sein könnten.
Wahrscheinlich waren Protozellen nur ca. 0,1 µm im Durchmesser, also etwa 10 bis 100fach kleiner als die meisten heutigen Zellen. Auf schematischen Darstellungen sind daher schon die einzelnen Atome und die daraus gebildeten Moleküle erkennbar. Das derzeit einfachste und bekannteste Modell für eine Protozelle besteht aus nur 2 Molekülklassen: Lipiden und Nukleotiden. Die Zellmembran wird aus Lipiden aufgebaut und im Inneren der Zelle befinden sich Polynukleotide (siehe Titelbild).
Die Hülle der Protozelle
Zur Klasse der Lipide gehört z.B. die Decansäure. Das Geheimnis eines solchen Moleküls ist, dass es aus zwei Teilen besteht, die sehr unterschiedliche Eigenschaften haben, aber durch eine chemische Bindung fest miteinander verknüpft sind. Der Säureteil ist sehr gut wassermischbar (wie Essigsäure), wohingegen der Decanteil nicht wassermischbar ist (wie Wachs). Gibt man solche Moleküle in Wasser, lagern sich diese selbstständig so zusammen, dass der wassermischbare Säureteil zum Wasser gerichtet ist und die nicht mischbaren Decanteile vom Wasser abgeschirmt werden. Es gibt verschiedene Gebilde, die diese Bedingung erfüllen. Es können kleine kompakte Kugeln sein (Mizellen) oder größere Zellen (wissenschaftlich Vesikel genannt), deren Wand aus einer Doppelschicht von Decansäure besteht. Der Säureteil der inneren Schicht zeigt dann nach Innen und der Säureteil der äußeren Schicht nach Außen, so dass sowohl im Inneren als auch Außen von der kugelförmigen Zelle Wasser ist. Genau aus solchen Doppellipidschichten sind auch heute alle Zellmembranen aufgebaut. Ob eher Mizellen oder Vesikel (Zellen) bevorzugt gebildet werden, hängt von der Sorte der Lipidmoleküle und den äußeren Bedingungen ab, z.B. dem Säuregehalt des Wassers.
Video: Bildung von Vesikeln aus Fettsäuremolekülen. Quelle: http://exploringorigins.org/resources.html; Illustration von Janet Iwasa, Lizenz Creative Commons Attribution-Noncommercial-No Derivative Works 3.0 United States License.
Wichtig zu erwähnen ist, dass die Lipidmoleküle sich nur lose zusammenlagern und nicht fest miteinander verbunden sind. Innerhalb der Zellmembran tauschen sie ihre Plätze, sind ständig in Bewegung und können auch zeitweise die Zelle verlassen und in die Zellmembran einer anderen Zelle eingebaut werden. Es handelt sich sozusagen um eine „flüssige“ Membran. Dadurch können kleine Nährstoffmoleküle in die Zelle eindringen. Alle diese Strukturen sind sehr klein und homogen im Wasser verteilt, so dass man sie mit dem Auge nicht sehen kann. Klar ist, dass die Bildung von Vesikeln ein normaler physikalischer Selbstorganisationsprozess ist, der keine separate Erbinformation benötigt, sondern rein durch die Eigenschaften der einzelnen Lipidmoleküle zu erklären ist. Dieser Prozess ist schon lange gut verstanden und wird in der chemischen Industrie aktiv ausgenutzt. Seife besteht ebenfalls aus solchen Molekülen und die Einlagerung von wasserunmischbarem Fett in die kugelförmigen Seifenmizellen ist dafür verantwortlich, dass wir nach dem Grillhähnchen essen wieder unsere Hände und Hemden sauber kriegen. Auch Seifenblasen lassen sich so erklären, nur dass noch Luft als dritte Komponente hinzukommt. Interessanterweise scheint die selbstständige Bildung von Seifenblasen weniger Verwunderung hervorzurufen als dies Selbstorganisationsphänomene im Kontext der Evolution tun.
Ungeklärt war lange Zeit, wie unter milden Bedingungen Zellwachstum und Zellteilung stattfinden können. Hierzu wurden in Harvard im Labor des Nobelpreisträgers Jack Szostak faszinierende Studien durchgeführt. Seine Gruppe konnte 2009 zeigen, wie Zellen wachsen, wenn neue Lipidmoleküle in die wässrige Lösung gegeben werden. Noch wichtiger war aber zu beobachten, dass bei weiterer Zugabe lange Filamente aus der Zelle wachsen und die gesamte Zelle schließlich aus verzweigten dicken Filamenten besteht. In diesem Zustand reichen leichte Erschütterungen um eine Teilung in mehrere kugelförmige Tochterzellen zu bewirken. [4] Ein solches System erfüllt somit schon zwei Bedingungen des Lebens. Es kann Nahrung aufnehmen und es kann sich reproduzieren. Schwer vorstellbar ist allerdings wie sich eine solche Zelle weiter entwickeln könnte, da der Fortpflanzungserfolg rein von den Umgebungsbedingungen, z.B. der Nahrungszufuhr, abhängt und in keiner Form in der Zelle kodiert und an die Nachkommen weitergegeben wird. Hier kommt die zweite Komponente der Protozelle ins Spiel: Die Polynukleotide.
Selbstreplizierende Erbinformation
Wie der Name andeutet, handelt es sich bei Polynukleotiden um eine Kette aus einzelnen Nukleotidbausteinen. Da es vier verschiedene Nukleotidbausteine in diesem Modell gibt, können sie auf verschiedene Weise zu Ketten verknüpft werden. Schon eine Kette aus nur 10 Nukleotiden kann auf 410, d.h. gut eine Million Arten, verknüpft werden. Die einzigartige Verknüpfungssequenz eines Polynukleotids stellt die Erbinformation dar. Durch das Studium zellulären Lebens wurden bereits zwei eng miteinander verwandte Polynukleotide gut erforscht, DNA und RNA. Beide verwenden vier Nukleotidbausteine, die molekulare Struktur dieser Bausteine unterscheidet sich jedoch leicht. Heutige Zellen verwenden DNA zum Speichern ihrer Erbinformation, wohingegen RNA zum Übertragen von Informationen und zur Beschleunigung von chemischen Reaktionen (Katalyse) eingesetzt wird. Die Entdeckung der Katalysefunktion durch Thomas Chech und Sidney Altman wurde 1989 durch einen Nobelpreis ausgezeichnet. Diese Doppelfunktion der RNA, sprich Speicherung von Erbinformation und Reaktionsbeschleunigung, ist auch von zentraler Bedeutung für das vorgeschlagene Modell einer selbstreplizierenden Erbinformation.
Und so funktioniert die Selbstreplikation: Ein Nukleotidmolekül besteht aus drei Teilen, einer Nukleinbase, Ribose und einem Phosphat. Im Polynukleotid sind die Phosphatteile mit dem Riboseteil des nächsten Nukleotids in der Kette verknüpft, so dass sich Ribose und Phosphat im Rückgrat der Kette befinden, wohingegen die Nukleinbasen seitlich abstehen. Die korrekte Verknüpfungsreaktion zwischen einzelnen Nukleotiden zur RNA läuft aber nicht ohne Hilfestellung ab. Die Phosphatgruppe und die Ribosegruppe müssen nah zusammengebracht werden um miteinander zu reagieren. Nun haben die seitlich von der RNA-Kette abstehenden Nukleinbasen noch eine besondere Eigenschaft. Sie bevorzugen die Anlagerung genau einer der vier Nukleinbasen. Dieser Prozess ist gut verstanden und folgt einem Schlüssel-Schloss Prinzip. Die vier Nukleinbasen werden mit dem Anfangsbuchstaben ihres Namens abgekürzt, A für Adenin, C für Cytidin, U für Uracil und G für Guanin. Die bevorzugten Paare sind GC und AU. Man nennt diese komplementär zueinander. Nehmen wir jetzt eine kurze RNA-Sequenz aus sechs Nukleotiden von CCGUAG an. Gibt man nun eine Mischung aller vier Nukleotide in die Lösung zu unserer RNA-Sequenz, so werden sich diese entsprechend an dieser „Mini-RNA-Kette“ anlagern, d.h. GGCAUC. Durch diese Anlagerung werden die Phosphatteile und Riboseteile der Nukleotide nah zusammengebracht und können sich leichter verknüpfen. So entsteht eine zweite RNA-Kette, die komplementär zur ersten ist. Bei etwas höheren Temperaturen können sich die beiden RNA-Ketten dann voneinander lösen. Das Interessante dabei ist, dass die neue komplementäre RNA-Kette natürlich auch als Vorlage für die Bildung der ursprünglichen RNA-Kette dienen kann. Somit enthält RNA nicht nur den Bauplan für eine komplementäre RNA, sondern im Prinzip auch den Bauplan für sich selbst. Nach diesem Prinzip könnten sich RNA-Ketten innerhalb von Zellen reproduzieren.
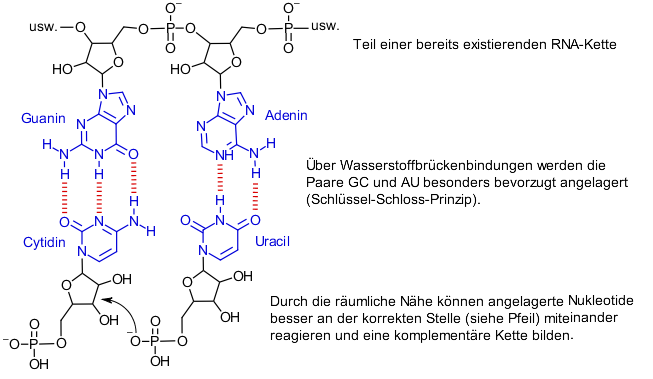
Der Haken an der Sache ist, dass die Reaktivität der Nukleotide auch mit diesem Anlagerungsprozess immer noch nicht ausreicht um effektiv längere RNA-Ketten zu bilden. Eine weitere Hilfestellung ist nötig und hier kommt die Doppelfunktion der RNA ins Spiel. RNA Ketten sind nicht steif sondern sind im Wasser wie eine Spaghetti flexibel. Schauen wir uns unsere erste Beispielsequenz CCGUAG noch einmal an, sieht man, dass die Cytidinbase am Anfang der Kette sich auch an die Guaninbase der selben RNA-Kette anlagern könnte, so dass sich ein Ring bildet. Hat man nun deutlich längere Sequenzen kann man sich vorstellen, dass sich viele solcher losen Querverbindungen durch Basenpaarung innerhalb der selben Kette bilden können, welche zu einer komplexen Knäuelbildung der „Spaghetti“ führen. In diesem Knäuelzustand kann RNA auch als Katalysator wirken (eine anschauliche schriftliche Beschreibung des Katalysemechanismus ist schwierig. Interessierte seien auf weitere Animationsvideos von Stated Clearly und dem Exploring Origins Project in englischer Sprache verwiesen. [15] [16]). Man nimmt also an, dass man für die Herstellung von RNA sowohl eine RNA-Kette als „Bauplan“ und Vorlage benötigt als auch eine „geknäuelte“ RNA Kette, welche bei der Verknüpfung der einzelnen Nukleotide hilft.
In der Tat hatte man 1996 RNAs gefunden, die in der Lage sind die Bildung anderer RNA-Sequenzen zu beschleunigen. [5] Die ersten Exemplare waren aber noch sehr ineffektiv und konnten nur ca. 10 Nukleotide miteinander verknüpfen. Solche kurze Sequenzen reichen aber nicht aus um ein wiederum katalytisch aktives „Knäuel“ bilden zu können. Man nimmt an, dass man hierzu mindestens 30-40 Einheiten benötigt. Außerdem hatte man lange Zeit das Problem, dass eine effiziente Verknüpfung nur an RNA-Ketten mit vielen C-Einheiten als Vorlage funktionierte. Forscher am Scripps Research Institute in Kalifornien konnten 2016 jedoch eine RNA-Sequenz finden, welche auch an C-ärmeren Vorlagen als Katalysator zur Bildung von über 70 Einheiten langen RNAs funktionierte, welche wiederum selbst als Katalysator für andere chemische Reaktionen funktionierten. [6] Den vollständigen Prozess der RNA-Replikation und Zellteilung kann man sich dann wie im folgenden Video vorstellen.
Video: Modellvorstellung von RNA-Replikation und Zellteilung. Quelle: http://exploringorigins.org/resources.html; Illustration von Janet Iwasa, Lizenz Creative Commons Attribution-Noncommercial-No Derivative Works 3.0 United States License.
Allerdings sind noch weitere Verbesserungen in Reaktionsgeschwindigkeit und Kompatibilität mit komplexen RNA-Sequenzen nötig, um eine vollständige Selbstreplikation zu ermöglichen. Zwar scheint dieses Ziel in greifbarer Nähe zu sein, aber wie schon einleitend erwähnt ist das Kriterium der Reproduktion nicht ausreichend für Leben.
Mutation: Die Evolution beginnt
Sehr selten kann es zu einer Fehlpaarung am Vorlage-RNA-Strang kommen. Anstatt GC, könnte eine GA-Paarung auftreten. Dadurch ist der neu gebildete RNA-Strang nicht mehr komplementär, sondern unterscheidet sich durch eine Punktmutation. In den meisten Fällen verliert die RNA dann ihre katalytische Funktion oder funktioniert schlechter als zuvor. In seltenen Fällen kann es aber vorkommen, dass die Mutation eine Beschleunigung der Selbstreplikation verursacht. Findet dieser Prozess in einer Zelle statt, kommt es in der Zelle zur Bildung einer erhöhten Anzahl von RNA-Ketten inklusive der verbesserten Variante. Teilt sich die Zelle in mehrere Tochterzellen ist es unwahrscheinlich, dass beide Varianten zu gleichen Teilen vorliegen, so dass sich die Tochterzelle mit der erhöhten Anzahl der besseren Variante mit größerem Erfolg gegen andere Zellen durchsetzen wird, denn: Bei zu langsamer RNA-Selbstreplikation kann es vorkommen, dass eine Tochterzelle nur eine einzige RNA-Kette enthält. Dies reicht nicht aus um den Selbstreplikationsprozess fortzusetzen und die Zelle stirbt. Nun wird auch die Bedeutung der Zelle für das Leben deutlich. Eine RNA mit verbesserter Selbstreplikation, die frei in Lösung herumschwimmt, würde schlechtere RNA-Varianten mit gleicher Wahrscheinlichkeit vervielfältigen. [7] Der Prozess der natürlichen Selektion funktioniert wesentlich besser bei räumlicher Trennung von verschiedener Erbinformation.
Nun sind auch Mutationen denkbar, die nicht direkt die Geschwindigkeit der Selbstreproduktion erhöhen, sondern die Geschwindigkeit anderer chemischer Reaktionen beschleunigen können, die der Zelle beim Überleben nützlich sind. Zum Beispiel könnten auch RNA-Ketten entstehen, die die Herstellung der Lipidmoleküle aus einfacheren Bausteinen ermöglichen. Dadurch könnte die Zelle auch in Umgebungen überleben, wo keine fertigen Lipidmoleküle vorliegen. Das gleiche ist denkbar für die vier Nukleotide. In späteren Evolutionsstufen könnten andere Sorten von Lipiden hergestellt werden, die zuvor gar nicht verfügbar waren, und helfen die Zellmembran stabiler zu machen. Die Synthese von Farbstoffmolekülen, welche sich in die Zellmembran einlagern um Sonnenlicht in Wärme umzuwandeln, könnte als zusätzliche Energiequelle für chemische Reaktionen dienen. Man sieht: es gibt unendlich viele Möglichkeiten um einen rapiden Komplexitätszuwachs zu erklären!
Die Protozelle ist sowohl zu Metabolismus und Reproduktion als auch zur Mutation fähig und erfüllt damit alle Kriterien für Leben. Die Entstehung der ersten Protozelle wirft aber weitere Fragen auf.
Woher kommt das erste Polynukleotid und wie kommt es in die Zelle?
Der aufmerksame Leser möchte vermutlich noch wissen, wie die Nukleotide und erst recht Polynukleotide, also die RNA, in die Zelle kommen. Auch hierfür gibt es plausible Erklärungen. Wie bereits erwähnt sind die Lipide in der Zellmembran nicht fest miteinander verknüpft, so dass sich die kleinen Nukleotidmoleküle durch die Zwischenräume schmuggeln können. Lange RNA-Ketten hingegen können die Zellmembran nicht durchdringen. Zu einem Einschluss von RNA in eine Zelle kann es kommen, wenn ein Gemisch aus Zellen und RNA eintrocknet, wobei sich die Zellmembran öffnet und Schichten bildet. Befindet sich eine RNA-Kette zwischen diesen Schichten kann diese bei erneutem Wasserkontakt in eine neugebildete Zelle eingeschlossen werden. [8]
Wenn die vollständige Selbstreplikation von RNA-Ketten aus den einzelnen Nukleotidbausteinen ausschließlich mit Hilfe einer besonderen katalytisch wirkenden RNA-Kette funktionieren kann, lässt sich nicht erklären wie die dafür mindestens notwendigen zwei RNA-Ketten entstanden sind. Aus diesem Grund muss dem oben beschrieben Mechanismus noch eine einfachere Möglichkeit der Selbstreplikation vorausgegangen sein. Hierfür wird angenommen, dass die Nukleotide in einer energiereicheren „aktivierten“ Form vorlagen, die sich aufgrund eines leicht abspaltbaren Teils (Abgangsgruppe) einfacher verknüpfen lässt. Für solche aktivierten Nukleotide konnte gezeigt werden wie lange zufällige Sequenzen von RNA an der Oberfläche von häufig vorkommenden Silikatmineralien entstehen können. [9] Die Möglichkeit der Selbstreplikation wiederum, d.h. die Entstehung von komplementären Ketten direkt an der bereits existierenden Vorlage, wurde in Wasser-Eis-Gemischen beobachtet. [10] Wenn Wasser anfängt zu gefrieren werden gelöste Bestandteile in der verbleibenden flüssigen Phase aufkonzentriert und können so einfacher miteinander reagieren.
Woher kommen Lipide und Nukleotide?
Schließlich bleibt natürlich die Frage wie die bisher genannten Grundbausteine, also Nukleotide und Lipide, entstanden sind. 1998 konnte gezeigt werden, dass sich Lipide, darunter auch die bereits genannte Decansäure, im Labor aus in Wasser gelöstem Kohlenstoffmonoxid und Wasserstoff unter hohem Druck und Temperatur bilden können. [11] Genau wie bei der RNA-Herstellung wurden hier gewöhnliche Silikatmineralien hinzugegeben an deren Oberfläche die Reaktionen stattfanden. Die beschriebenen Reaktionsbedingen für die Lipidsynthese sind charakteristisch für hydrothermale Quellen wie beispielsweise Geysire. Allerdings entstehen die Lipide hier in zu kleinen Konzentrationen um sich zu Mizellen zusammenzufinden. Bricht der Geysir aus, könnten Wassertröpfchen mit Lipiden über weite Strecken transportiert werden. Unterwegs oder am Zielort können diese durch Verdunstung aufkonzentriert werden, so dass schließlich Mizellen entstehen.
Einer der größten Forschungserfolge im letzten Jahrzehnt konnte bei der Nukleotidsynthese gefeiert werden. Die Entstehung der Nukleotide unter den primitiven Bedingungen der Urerde wurde lange Zeit als Unmöglich betrachtet und als Hauptkritikpunkt gegen die in diesem Artikel beschriebene Protozellenhypothese angeführt. Wie schon erwähnt besteht ein Nukleotidmolekül aus drei Teilen. Der entscheidende Teil für die Unterscheidung der vier in der RNA vorkommenden Nukleotide ist der Teil der Nukleinbase (Adenin, Cytidin, Uracil und Guanin). Der Ribose- und Phosphatteil sind bei allen identisch. Alle Teile sind auch als eigenständige Moleküle gut bekannt und um den Aufbau der RNA zu beschreiben ist diese Unterteilung nützlich. Diese tradierte Sichtweise hatte allerdings zur Folge, dass man lange vergeblich versuchte die Nukleotide aus eben diesen drei Teilen herzustellen. Erstens existierte kein plausibler Weg die Nukleinbasen alleine in vernünftigen Ausbeuten herzustellen, zweitens ist der Riboseteil alleine extrem instabil und drittens gelang es nie in zufriedenstellender Weise Ribose und die Nukleinbase an der richtigen Stelle miteinander zu verknüpfen. [12] 2009 gelang es dem Doktoranden Matthew Powner in der Arbeitsgruppe von Jack Szostak erstmalig eine plausible Syntheseroute für die Cytidin- und Uracil-haltigen Nukleotide zu zeigen. [13] Das Faszinierende daran ist, dass die Synthese erstens mit einfachsten Rohstoffen startet, die keiner weiteren Rechtfertigung bedürfen, und zweitens, dass weder freie Ribose, noch die freie Nukleinbase in einem der Reaktionsschritte auftauchen. Ein plausibler Mechanismus für die Entstehung aller vier Nukleotide, d.h. auch der Adenin- und Guanin-haltigen Nukleotide, wurde noch nicht gefunden, aber man hofft, durch ähnliche Perspektivwechsel einen Weg zu finden. Mittlerweile sind auch deutsche Forscher wie Prof. Thomas Carell von der LMU München mit wichtigen Beiträgen am Rennen um den Durchbruch beteiligt. [14] Man drückt die Daumen.
So what?
Der in diesem Artikel skizzierte Mechanismus wird derzeit in der wissenschaftlichen Community als am Wahrscheinlichsten eingestuft. Natürlich gibt es zahlreiche alternative oder ergänzende interessante Modelle, wie z.B. selbst-replizierende Peptide oder die Entstehung des Lebens außerhalb der Erde (Panspermia-Hypothese). Diese Überlegungen stehen im gesunden wissenschaftlichen Wettstreit miteinander und sollen hier nicht diskreditiert werden. Vielmehr geht es darum, dem Leser zu verdeutlichen, dass eine kohärente naturwissenschaftliche Lösung dieser Fragestellung sehr gut vorstellbar ist und mit an Sicherheit grenzender Wahrscheinlichkeit auch gefunden wird.
1543 publizierte Kopernikus seine Theorie vom Heliozentrismus. Damit stand die Erde nicht mehr im Mittelpunkt unseres Sonnensystems. Giordano Bruno spekulierte 1584, dass die Sterne andere Sonnen sein könnten, die ebenfalls Planeten besitzen. Er wurde daraufhin von der Inquisition 1600 auf dem Scheiterhaufen verbrannt, aber seine Hypothesen wurden von späteren Astronomen bestätigt. Nun war auch die Sonne nicht mehr Mittelpunkt des Universums. 1859 publizierte Charles Darwin „The Origin of Species“. Es wurde klar, dass der Mensch eine Trockennasenaffenart ist und nicht etwa die Krone der Schöpfung. All diese Erkenntnisse wurden zunächst als Kränkung empfunden und hatten es daher (und haben es teilweise noch immer!) schwer akzeptiert zu werden. Wir sollten uns darauf vorbereiten, dass die Entschlüsselung des Ursprungs des Lebens eine vergleichbar heilsame Revolution unseres Selbstverständnisses zur Folge haben wird.
Wer gerne eine pdf-Version ohne Videos dieses Artikels hat: Suetterlin_Chemische_Evolution_Forschungsstand_2018
Danksagung
Für kritische Hinweise und Verbesserungsvorschläge möchte sich der Autor bedanken bei Dirk Brixius, Dr. Michael Salz, Till von Reumont, Tobias Stelmaszyk, Dr. Joachim Reimer und Dr. Bernhard Seuring. Dem „Exploring Origins Project“ des Museums of Science (Boston, USA) in Zusammenarbeit mit der Arbeitsgruppe von Jack Szostak (Harvard Medical School) ist zu Danken für die Erstellung der großartigen und vor allem offen lizensierten Grafiken und Videos (Janet Iwasa).
Literaturverzeichnis
| [1] | А. И. Опарин, Произхождение жизни, Москва: М. МОСКОВСКИЙ РАБОКИ, 1924. |
| [2] | A. I. Oparin, Die Entstehung des Lebens auf der Erde, Berlin: Volk und Wissen Verlag Berlin-Leipzig, 1949, p. 245. |
| [3] | H. Rauchfuß, Chemische Evolution und der Ursprung des Lebens, Berlin Heidelberg: Springer, 2005. |
| [4] | T. F. Zhu und J. W. Szostak, „Coupled growth and division of model protocell membranes,“ Journal of the American Chemical Society, Bd. 131, Nr. 15, pp. 5705-5713, 2009. |
| [5] | E. H. Ekland und D. P. Bartel, „RNA-catalysed RNA polymerization using nucleoside triphosphates,“ Nature, Bd. 382, pp. 373-376, 1996. |
| [6] | D. P. Horning und G. F. Joyce, „Amplification of RNA by an RNA polymerase ribozyme,“ Proceedings of the National Academy of Sciences, Bd. 113, Nr. 35, pp. 9786-9791, 2016. |
| [7] | J. W. Szostak, D. P. Bartel und L. Luisi, „Synthesizing Life,“ Nature, Bd. 409, pp. 387-390, 18 January 2001. |
| [8] | D. W. Deamer, „Membrane Compartments in Prebiotic Evolution,“ in s The Molecular Origins of Life, Cambridge, Cambridge University Press, 1998, p. 189. |
| [9] | J. P. Ferris, „Montmorillonite-catalysed formation of RNA oligomers: the possible role of catalysis in the origins of life,“ Philosophical Transactions of the Royal Society B, Bd. 361, pp. 1777-1786, 29 October 2006. |
| [10] | J. Attwater, A. Wochner, V. B. Pinheiro, A. Coulson und P. Holliger, „Ice as Protecellular Medium for RNA replication,“ Nature Communications, Bd. 1, pp. 1-9, 21 September 2010. |
| [11] | M. M. Thomas, G. Ritter und B. R. T. Simoneit, „Lipid Synthesis Under Hydrothermal Conditions by Fischer- Tropsch-Type Reactions,“ Origins of life and evolution of the biosphere, Bd. 29, pp. 153-166, March 1999. |
| [12] | S. Islam und M. W. Powner, „Prebiotic Systems Chemistry: Complexity Overcoming Clutter,“ Chem, Bd. 2, Nr. 4, pp. 470-501, 13 April 2017. |
| [13] | M. W. Powner, B. Gerland und J. D. Sutherland, „Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions,“ Nature, Bd. 459, pp. 239-242, 2009. |
| [14] | S. Becker, I. Thoma, A. Deutsch, T. Gehrke, P. Mayer, H. Zipse und T. Carell, „A high-yielding, strictly regioselective prebiotic purine nucleoside formation pathway,“ Science, Bd. 352, Nr. 6287, pp. 833-836, 13 May 2016. |
| [15] | „What Is the RNA World Hypothesis?,“ Stated Clearly, August 2016. [Online]. Available: http://statedclearly.com/videos/RNA-World/. [Zugriff am 9. Juni 2018]. |
| [16] | „Replicase ribozyme movie,“ [Online]. Available: http://exploringorigins.org/resources.html. [Zugriff am 9. Juni 2018]. |